geographical location of
the peat pond
Pond Bergknappweiher (47°51’9.99''N,
11°14’18''E) is situated in the pre-alpine
region in
Bavaria
(Germany), about
50 km southwest of the city of
Munich. The
pond is
very close to the both lakes Ammersee
S
and Starnberger See, about
15 km
and 6 km away, respectively.
This pond was created by peat cutting.
the dystrophic character and seasonal
phytoplankton development
 Cyanobacteria in
Bergknappweiher, 2001:
Cyanobacteria in
Bergknappweiher, 2001:
The cyanobacterial surface scum of this dystrophic pond in autumn is
mainly composed of bundles of Aphanizomenon
flos-aquae but also of various
Microcystis and few Anabaena taxa.
The photos in the gallery illustrate the
appearance
throughout the
seasons of Bergknappweiher. It is a dystrophic
pond, which means
that
the water contains high
concentrations of dissolved organic carbon
(DOC). DOC makes water looking yellow-brownish (see photo
8 in the
above gallery). The colour of
the water, or more specifically the high concentrations of
DOC, is, however, harmless for bathers and life in the peat pond. It
could even be seen as a positive environmental factor
as a water body with high DOC content filters out much stronger UV in
comparison with a pond or lake with clear water. For example, in a
clear water lake in the alpine region (low DOC concentration),
Lake Lucerne
at 454 m above sea level in the Swiss Alps, UV
significantly inhibits the photosynthesis of primary producers within
the top
5 m on a sunny day (see Fig.2
in Teubner et al.
2001 R).
Measuring photosynthesis at water surface at 0 and 2 m
depth in lake Lucerne, the water samples that were shielded from
the
UV-A-part of the spectrum, yielded much higher
photosynthetic rates
than those samples at the same depth but without UV-protection. It
could be further seen from the vertical profile of photosynthetic rates
that the inhibition effect by UV decreases with depths below
2 m. At a
depth of about 5m the photosynthetic rates of UV-protected and
non-protected samples were in the same range. Pond Bergknappweiher is
located in the pre-alpine region at about 617 m above sea
level, but DOC
might shield primary producers from UV even at the top surface of pond
water. The aquatic life in water basins in the alpine region can become
more vulnerable against UV in case the water basins are at high
altitude and of low DOC concentration.
In Bergknappweiher, the high concentration of DOC is due to ‘old’
humic
substances that are naturally occurring as organic
components from the
soil (‘fresh/young’ DOC in a water for example can be released by algae
being alive in the water body). The pond was created by peat cutting
and is still surrounded by meadows and also woodland. The meadows are
today mainly used as pastures for cattle or haymaking. Further details
about
sustainable animal husbandry in this pre-alpine region are
illustrated on the website about Ammersee
S,
for the alpine region in the
Salzkammergut district in Austria on the website about Ammersee
S.
 Bergknappweiher, 2001:
Bergknappweiher, 2001:
Meadows and woodland are surrounding the pond.
.
 Bergknappweiher, 2001:
Bergknappweiher, 2001:
Tufted forms of large Carex
species (Carex
acutiformis/riparia,
here seen as still small plants in spring, are found at the pond’s
edge.
 Along the shore
of Bergknappweiher, 2001:
Along the shore
of Bergknappweiher, 2001:
This photo is taken on the road going along the shore of the pond.
Sustainable animal husbandry in this pre-alpine region in Bavaria
(Germany) is in a more detail
illustrated on this website about Ammersee
S.
See also sustainable agriculture in the alpine region in Upper Austria
on the website about Attersee S.
 Landscape nearby Bergknappweiher, 2001:
Landscape nearby Bergknappweiher, 2001:
Small ponds created by peat cutting are quite common in this area. The
pond seen on the photo is in the close neighbourhood of the described
pond Bergknappweiher.
Plants grow, bloom and built-up fruits and thus change the
appearance
of the meadows and woodland in the course of a year. However, not only
communities of plants and animals living around a water basin change
with seasons, but also the communities of aquatic microorganisms
inhabiting the basin do so. The
seasonal changes are usually not by
chance but follow a pattern driven by the changing environment
throughout the seasonal cycle. The development of phytoplankton in pond
Bergknappweiher will be illustrated in a greater detail in the
following paragraph, as the seasonal pattern described here can be
typically found in eutrophied small basins of stagnant water.
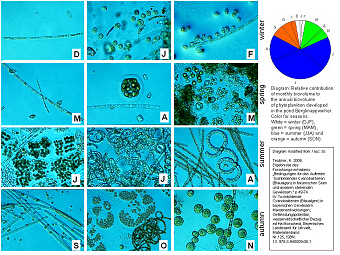 Phytoplankton of
Bergknappweiher,
2000/2001:
Phytoplankton of
Bergknappweiher,
2000/2001:
Seasonal development of phytoplankton biovolume observed from December
2000 to November 2001. The DIC-photos are from light microscopy (DIC =
differential interference contrast). The graph
on the right side shows the relative contribution of monthly biovolume
to the annual biovolume of phytoplankton for the studied period. The
seasons are indicated in white for winter, green for spring, blue for
summer and orange for autumn.
The figure on the left side illustrates the monthly
shifts among
phytoplankton species and phytoplankton biovolume throughout the year.
The lowest phytoplankton biovolume is estimated
for the wintertime
(see white labelled area in the circular graph).
Some diatoms (e.g. Aulacoseira
as seen for winter on the microscopical photo leftside)
and chlorophytes are
found in the water column during the cold season. With increasing day
length, the phytoplankton cells are growing and build a spring peak of
biovolume. With regard to the chilly period from early autumn to
late spring (September to May), highest biovolumes can be developed
in March, as there are sufficient nutrients available for an unlimited
growth. The nutrients were accumulated over the cold season. They are
now well distributed along the water column due to spring turnover
(mixing)
and can be thus utilized by
phytoplankton organisms floating in the water body. During this
period early in the year the exponential
growth of fast growing
small-sized single-cell phytoplankton species such as
small
centric diatoms,
needle-shaped diatoms and small cryptophytes and chlorophytes
predominates.
The dominance of small species in spring can be identified by peak
values of the cell-surface to cell-volume ratio of phytoplankton (see
dynamic of this ratio in riverine lakes described on the website of Grosser Mueggelsee
S).
A few weeks later, in April, the biovolume is again
remarkable low. It is almost the lowest for the whole year in
Bergknappweiher
as seen in the circular diagram. This weak phytoplankton biovolume is
due to grazing of zooplankton.
The growth of zooplankton depends more
on temperature than photosynthetic organisms, i.e. phytoplankton
organisms. As there is a time shift between the increase of day length
and an increase of WATER temperature (see Mondsee
S
and Ammersee
S
about
coherence and time-shifts between annual peaks of temperature related
parameters referring to lake physics and lake biota), a time-lag
between phytoplankton and zooplankton species can be often seen. After
the clearance of phytoplankton cells the numbers of zooplankton
organisms are also lowering as no further food is available anymore.
This short
interim scenario of a low number of biotic particles in the water
column (almost no phytoplankton and also no zooplankton) is reflected
by a temporarily high water transparency lasting few days to a week and
is called ‘clear water phase’.
In May, the phytoplankton species
are growing well again even if the species composition is quite
different
than in early spring. The phytoplankton composition is now dominated by
large diatoms and large colonial green algae (Coenochloris
spec.).
During summer,
i.e. the period from June to August, about 70% of
the annual phytoplankton biovolume are built up in Bergknappweiher as
indicated by the large blue area in the circle diagram. The month with
the highest biovolume development here is July. The summer biovolume is
mainly built up by cyanobacteria. It might be worth emphasizing that
the
contribution of species to higher taxa as e.g. the cyanobacteria may
change
drastically from month to month. In June, some chroococcale
cyanobacteria as Microcystis
(among other M. viridis)
are predominant
while in July and August filamentous cyanobacterial forms as Anabaena spp. and Aphanizomenon
flos-aquae are
common in Bergknappweiher. Many
of the cyanobacteria seen on these photos are known to produce toxins.
The summer cyanobacteria in Bergknappweiher built large colonies and
are usually not the preferred food for zooplankton for many reasons.
Due to the shallowness of the pond, the water is well mixed throughout
summer. Consequently, autumnal turnover is not important to replenish
the
nutrients from the deep near-sediment zone and hence no further peak,
i.e. no autumnal phytoplankton peak, can be expected. This contrasts
with the situation in spring where the biovolume development did not
succeed evenly month by
month, the phytoplankton biovolume in autumn
lowers successively. Cyanobacterial
forms that were dominant in late summer persist throughout autumn (see
also summer/autumn and winter/spring phytoplankton composition
described for riverine lake on the website Grosser
Mueggelsee
S).
As seen
on the microscopical photomicrograph for the September sample,
Aphanizomenon
flos-aquae already forms dormancy stages to survive well during the
non-growing season. Bergknappweiher provides an example for dynamic
phytoplankton development in terms of
biovolume evolvement and species composition throughout the year. How
many samples at what time in the year should be taken to get reliable
data
assessing such an ecosystem? Is one sample a year sufficient
or do we need to spend more effort and need to take samples two or four
or six times a year to get a reliable data set to assess such a water
body? Scientists and government agencies, and also landscape planners
constructing swimming pools, need to answer such questions about
the schedule of useful sampling
intervals. Some aspects of taking
samples in a deep lake with a deep chlorophyll layer are discussed on
the website about Mondsee
S.
Some further background about general
pattern of seasonal development of phytoplankton and of nutrient
availability is described on the websites about Grosser
Mueggelsee
S.
In summary,
Bergknappweiher provides an
example of a shallow water body, which is covered by a
cyanobacterial
scum on calm days during the growing season. Many
cyanobacteria
are present on the top water surface, giving the water in
particular a greenish hue. Cyanobacterial scum or blooms in stagnant
water
bodies are not a phenomenon of a certain country or region but occurs
worldwide. Cyanobacterial blooms are often associated with nutrient
enrichment of the water body. They are described on this website for
shallow lakes as Old Danube
S
, Grosser Mueggelsee
S
& Langer See
S,
Taihu
S,
Poyang
S and
other ponds
such as
Biotop Auersthal
S.
Cyanobacteria, hoewever, can be also abundant in deep lakes
such as Ammersee
S
and Mondsee
S.
Some
further aspects about the seasonal development of
phytoplankton in shallow lakes are described on the page about Grosser Mueggelsee
S.
 >
>