geographical location of the
urban lake
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
North basin (Upper Old Danube) with a view in north direction over the deepest site
towards the entrance of the lido Angelibad. The deepest site of the Old Danube, the
"Birnerloch", is 7m deep. The mean depth is 2.5m.
Old Danube (48°14'9.26''N,
16°25'41.6''E)
is a shallow urban lake in the
city Vienna
that was cut off
from the
main river Danube about 160 years ago during river
regulation (see map Fig.1
in Dokulil et al.
2000 R,
Fig.1
in Donabaum et al.
2004 R).
This groundwater-seepage
lake has a water volume of 3.7 x 106
m3, an area of 1.5 km2
and maximum depth of 6.8m. The theoretical water retention time was
about 190 days before water level drawdown in recent years. The oxbow
lake is still a popular
recreational area and attracts people for boating,
swimming or just for walking around the lake in the capital of Austria. Paying attention to
this high recreational value, the City of Vienna has taken measures to improve and later to
maintain the water quality in the Old Danube for many years
(Dokulil, Donabaum and Teubner 2018 R,
Teubner et al. 2020 R,
2022 R).
In 1987, underwater plants and Chara-beds covered almost the
whole water basin of Alte Donau (Fig.4
in Donabaum et al.
2004 R).
The pre-dominance of submerged
macrophyte growth was at the expense of the growth of photosynthetic
microorganisms drifting in the water (Fig.4
in Dokulil et al.
2000 R).
During this stable state of
macrophyte dominance,
the water transparency was high (Teubner et al., 2020 R,
2022 R). Old Danube, however,
underwent large
ecosystem changes. The nutrient
input, mainly phosphate, increased markedly in 1988 and
1989 and slightly further until 1994. At the same time a loss of 95% of
macrophyte biomass but an increase of biomass of drifting
photosynthetic microorganisms in the water column, namely formation of
heavy cyanobacterial blooms, took place. The main cyanobacterium was at
that time
Cylindrospermopsis
raciborskii (Figs.4&5
in Dokulil & Teubner 2000 R).
It contributed up to 63% of the
total biovolume
of drifting phototrophic organisms, i.e. phytoplankton organisms, in
Alte Donau (page 1143
in Teubner et al.
2003 R).
 Oxbow lake Old Danube /
Alte Donau, 2000:
Oxbow lake Old Danube /
Alte Donau, 2000:
View onto a small part of the south basin (Lower Old Danube). The island
"Gänsehäufel" is seen on the right side. The island's name refers to a "heap for
geese", two alluvial islands that were formed before the cut-off from the main river
bed in 1870 and which may have been used for geese breeding.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
View onto the small basin "Kaiserwasser", which is connected to the southern basin
(Lower Old Danube).
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
View along the western branch of the two arms of the southern basin (Lower Old
Danube).
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
View of a small part of the northern basin (Upper Old Danube), near the subway
station of the U1.
Lake
restoration in Old Danube focused primarily on the reduction of
phosphorus
(Figs.1&2
in Teubner et al.
2003 R),
as this is known to be the most limiting nutrient
element for phytoplankton growth in many freshwaters .
In the
following
years other lake treatments, mainly bio-manipulation,
have been moved on. A major effort was devoted to the resettlement of macrophytes
to the
lake
(for further information about macrophyte management in Old Danube
contact SYSTEMA).
The methods
for assessing the water quality by aquatic macrophytes acc.
to European Water Framework Directive are described in detail by Pall
&
Moser (2009 R).
The early recovery from eutrophication during an early stage of
restoration is briefly described in Donabaum et al. (2004 R).
The ecosystem change during sustainable management of lake restoration, concerning abiotic
and in particular many biotic aspects (from planktonic bacteria and ciliates, phytoplankton
and macrozoobenthos to fish and water plants), are summarized for more than 20 years for
Alte Donau in Dokulil, Donabaum and Teubner
(2018, R ).
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
South end of the south basin near the gauge "Seestern", Lower Old
Danube.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
North end of the Upper Old Danube, view into the north appendix
"Wasserpark".
The long-term
study of the lake
restoration of the Old Danube
covers four periods: (1) the situation of the nutrient-rich status in 1993-94
before the restoration started, (2) the following two years during the chemical treatment of
phosphate precipitation (phosphate flocculation by RIPLOX-treatment, period 1995-96) and
another three years of further restoration management (1997-1999), (3) reintroduction of
macrophytes by periodical water level draw down (2000-2006, further information about
macrophyte management contact SYSTEMA) and from 2007 onward the period of
ecologically stable conditions (further information about the lake restoration of the Old
Danube DWS-Hydrologie-Oekologie).
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
The submerged bushy water plant with reddish stems and rosettes of leafs can easily
be seen in the clear water. This water plant Spiked
Water-milfoil (Myriophyllum spicatum) grows well
utilizing the nutrients in the Old Danube and thus limits the nutrient availability
for unwanted photosynthetic microorganisms as the planktonic algae.
The submerged plants thus maintain the high water transparency in the Old Danube
(Teubner et al., 2020 R,
2022 R). In addition, their
dense stands in the littoral zone serve as valuable habitats for fish and other
animals.
 Oxbow lake Old Danube /
Alte Donau, 2013:
Oxbow lake Old Danube /
Alte Donau, 2013:
After the successful resettlement of submerged vegetation in the whole
lake, which improved the water quality (Teubner et al., 2020 R,
2022 R), the growth of these
plants needs to
be restricted in certain areas of the lake. This underwater-weed
harvester is used to keep such plants (submerged macrophytes) away from
the main shipping route and bathing
areas.
It is worth
mentioning that a drastic reduction of the nutrient load is best
achieved by an effective sustainable restoration management in the
catchment of a lake, in advance to an expensive in-lake restoration
treatment as described here for the Riplox technique below. It further
has to
be said, that a lake reverts to a good water quality, back
to its 'natural' reference status after a sustained reduction of the
nutrient load from the catchment. In this way it is less
expensive than an active in-lake restoration treatment, even if it
typically takes a few years longer. After passing the turning point due
to a
successful lake catchment restoration, the ecosystem follows a trend of
consecutively diminishing nutrient availability over years.
The expected time needed to accomplish a turn back to the ‘lake
reference status’ depends on the extent of water pollution, size of
the lake and the
water retention time among others, but would be about
4 to 8 (or even
12) years, which relates to a period of one to two (or even
three)
legislative sessions in many counties. Both sides of an ecosystem
response to
nutrient changes, the increase eutrophication
or the consequent reduction of nutrients (re-oligotrophication or more
accurately
to say oligotrophication),
do not happen ‘overnight’ but proceed over many seasonal cycles, over
years, usually with a delayed response due to an ecosystem
hysteresis.
The delayed response of phytoplankton to
oligotrophication has been described on this website for the deep
alpine lake
Mondsee S
( see also page 2 and Fig.4
in Dokulil & Teubner
2005 R).
Whatever the restoration of a lake is - only by reduced nutrient load
in the catchment or in addition by in-lake methods; the information
about the progress of oligotrophication is needed and hence an
advanced
research is appropriate to understand individual processes and trends
during the
ecosystem change.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
The freshwater jellyfish (Craspedacusta sowerbii) is
temporarily found during hot summer periods in some years in the Old Danube R. This limnomedusa can
easily be overlooked in the water column.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
Two freshwater jellyfish (Craspedacusta sowerbii) and
the water plant Spiked Water-milfoil (Myriophyllum
spicatum) taken into a bowl of water.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
Size comparison between the freshwater jellyfish (Craspedacusta sowerbii) R
and the water plant Spiked Water-milfoil (Myriophyllum
spicatum).
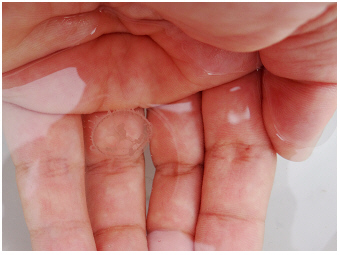 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
Tiny limnomedusa (Craspedacusta sowerbii) on a hand.
A note about this freshwater jellyfish from the perspective of divers is described
in Mathä (2009); the species description as "neobiota", their widespread
distribution in freshwater habitats and phylogeny is specified in Fritz et al.
(2007, references see list below).
the implementation of the RIPLOX-method
to reduce phosphate availability for algal growth by phosphate
flocculation in old danube
The Riplox-method
(Ripl 1976, R)
was chosen as the main internal restoration method to
reduce phosphate availability for phytoplankton growth. The method acronym Riplox is
according to the family name of Prof. Wilhelm RIPL, who
described the
lake restoration technique of OXidizing
the sediment surface to prevent internal nutrient loading by the
phosphate release from
anaerobic
sediments in heavily eutrophied lakes. Due to
the costs for in-lake restoration, this technique is usually applied
to smaller, rather than to larger water basins.
In Old Danube, the Riplox-restoration method was implemented
twice, in the spring of 1995 and
1996 (scheme of RIPLOX-treatment in Fig.6
in Dokulil et al.
2000 R).
Two steps were carried out on each occasion: First, FeCl3
buffered with limestone, was added to the water surface to
remove phosphorus and suspended material by chemical and mechanical
flocculation. After this, Ca(NO3)2
was added to the sediments to enhance nitrate oxidation. The mode of
RIPLOX-method is the following: With enhanced nitrate
oxidation, nitrate is then reduced to elemental nitrogen by anaerobic
denitrification. Organic mud is hence oxidized to carbon-dioxide and
water. Consequently, the oxygen deficit that is caused by heterotrophic
metabolism is lowered. Thus, prolonged anoxic conditions at the
sediment surface which is common in a nutrient-rich lake can be
prevented. The
internal phosphate loading by release from the sediment is thus
avoided. This initial treatment in Old Danube indeed shifted the
pelagic community, i.e. the micro-biota drifting in the free water
zone, from nutrient-rich conditions in 1994 to a sustained lower
nutrient status in 1995/1996 as described by the decrease of phosphorus
concentration and the response by the microbial community in detail
below.
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
The lake restoration included the management of the littoral zones. This replanted
littoral zone at the land-water interface of "Kaiserwasser" refers to the natural
zonation of vegetation in ecologically intact lakes: the landward Phragmites-reed belt is continued with the Typha reed-belt in the shallow littoral followed by the
floating leaf zone of submerged macrophytes towards the open water.
 Oxbow lake Old Danube / Alte Donau, 2015:
Oxbow lake Old Danube / Alte Donau, 2015:
The Greater Bladderwort (Utricularia vulgaris) was
found in the floating leaf zone of submerged vegetation shown on the left photo. The
small yellow flowers of this rather rare aquatic plant can easily be overlooked
nearby the dense stands of the Spiked Water-milfoil (Myriophyllum spicatum).
 Oxbow lake Old Danube /
Alte Donau, 2015:
Oxbow lake Old Danube /
Alte Donau, 2015:
The reconstructed littoral zone on the east shore of the eastern branch of the two
arms of the southern basin serves well as habitat for many animals. A frog, for
example, is seen on the leftside of the photo, close to the stands of Common Reed
(Phragmites australis).
 Oxbow lake Old Danube / Alte Donau, 2015:
Oxbow lake Old Danube / Alte Donau, 2015:
This photo is taken at the same place as the photo on the left-side and shows a frog
(Rana esculenta) in detail.
reduction of total phosphorus: ecosystem
assessment rather by concentrations of total
phosphorus than of phosphate
Phosphorus is known
as the nutrient
element mainly responsible for limiting the growth of algae and
cyanobacteria in many
freshwaters and also some marine systems. Phosphate
dissolved in the water is the only chemical phosphorus species that can
be utilized immediately by photosynthetic organisms as a nutrient
source. A further phosphorus fraction, the dissolved organic phosphorus
can only be utilized via extracellular enzymes released by bacteria and
primary producers. The fraction of bound phosphorus in the living
microorganism cells, i.e. the particulate organic phosphorus, is a
suitable nutrient source for algal growth NOT at all. For these reasons
dissolved inorganic phosphate indeed is used as the nutrient source for
phosphorus uptake
experiments. Such empirical phosphate additions are used
to conduct
algal
culture experiments in the laboratory or bioassay
fertilizing studies with phytoplankton in a lake.
These
experiments may aim at measuring the short-term
course of phosphorus acquisition of algae in the lab, or
the bioassay outgrow
experiments in a longer run over days identifying
potential algal species shift among phytoplankton in the lake.
 Oxbow lake Old Danube /
Alte Donau, 2000:
Oxbow lake Old Danube /
Alte Donau, 2000:
This photo, taken at the winter-time after the chemical RIPLOX-treatment of lake
restoration, shows a
high water transparency. Water transparency in winter, however, is not
indicative for the trophic state of an ecosystem (Teubner et al., 2020 R). The low turbidity in
winter might not be necessarily due to a low nutrient availability and,
hence, does not refer to nutrient-limited growth of the micro-organisms floating in
the
water body
(phytoplankton). It is simply due to the low temperature and short day
length that many phytoplankton species cannot develop a biomass peak as
usually achieved during the growing season. In lakes of the
temperate zone, the phytoplankton biomass usually peaks two or even three times a
year, in late spring and mid
summer, and in some lakes also in early autumn.
Phosphate, however, appears not to be an
accurate indicator of phosphorus resources
in a lake or river
ecosystem. The analytical detection limit for phosphate, measured as soluble reactive phosphorus
(SRP), depends on the method and is usually about 60-140 nano mol L-1.
In the case where the phosphate concentration in lake water is below
the analytical detection limit, it does not necessarily mean that ‘no’
phosphate is
available for growth and consequently ‘no’ algal biomass may increase.
This evidence comes from field studies. It can be shown that during
periods when phosphate in the epilimnetic lake sample is below the
analytical detection limit for days to weeks, the biomass of algae and
cyanobacteria may even increase at the same time. One reason is that primary producers are able to utilize
phosphate within minutes even below the concentration
range of the analytical detection limit (Teubner et al., in
preparation). A second reason can be seen in
the patchy environment as larger phosphate concentrations are indeed
available but only at very small dimensions of a few millimeters as by
point sources provided by zooplankton excretion (Teubner et al., in
preparation). Such patchy
ephemeral phosphate sources
also seem to be utilized by algae (see page
Preface S
). A third reason might be seen in
the
particular short turnover
time for phosphorus ranging from hours to days. Bound
phosphorus in biota or dissolved organic forms is after short microbial
cycles again available as a dissolved form of phosphate and can
be rapidly utilized. A last reason why phosphate concentration
measures in a
lake or river are often ‘zero’ even at periods of phytoplankton biomass
increase is that regular sampling is usually done during the daytime.
Phosphate
concentrations often varies with the day-night
cycle and are frequently, for some ecological reasons,
considerably higher during the night than daytime (Teubner et al., in
preparation). When phosphate is
limiting the growth of phytoplankton in a lake or river, the night-time
released phosphate can be day-time rapidly incorporated by biota.
Therefore, after enhanced concentrations during the night, the
phosphate can be again within minutes to few hours below the analytical
detection limit at day-time (Teubner et al., in preparation). To meet
these
four concerns, the total
phosphorus concentration
(TP) is usually measured to assess the nutrient state of an aquatic
ecosystem (examples for the Old Danube: Fig.4
in Dokulil et al.
2000 R,
Fig.5
in Donabaum et al.
2004 R,
examples for other lakes on this website see Mondsee S, Traunsee S
and
Taihu S).
In the case study of Old Danube restoration, the concentration of total
phosphorus (TP)
decreased drastically from 2.4 in 1994 (before treatment) to 0.6 µmol L-1
in 1996 (after treatment by RIPLOX-phosphate flocculation) in the water
body. According to ecosystem nutrient-state phytoplankton-response
curves, where concentrations of total phosphorus are plotted versus
concentrations of chlorophyll-a (this pigment that is used as the rough
estimator for biomass of primary producers), Old Danube changed
from a hypertrophic
to a moderately eutrophic ecosystem.
microbial community response to reduced
phosphate availability:
shift toward phosphorus-rich biota under
phosphorus-deficient conditions
 Oxbow lake Old Danube /
Alte Donau, 2011:
Oxbow lake Old Danube /
Alte Donau, 2011:
Taking a phytoplankton sample in spring for practical algal
courses at the University of Vienna. The species composition will be identified
from
a life sample and a fixed water sample under the microscope later in
the lab.
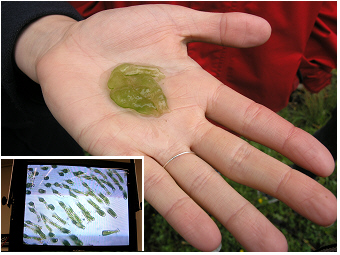 Oxbow lake Old Danube / Alte Donau, 2011:
Oxbow lake Old Danube / Alte Donau, 2011:
This 'green jelly colony' is found in spring at the shallow littoral
zone of the
reed
belt of the lake. Although it looks yellow-green, it is primarily not a alga
but an
animal, Ophrydium
versatile.
The insert shows the detail of this live colony under the
microscope: the periodicly contracting and expanding cells are
unicellular
animals,
ciliates. The green color is caused by endosymbiontic
green algal cells, of zoochlorellae, that are living inside these
protozoans. This species is common in lakes with a moderate
nutrient level and should not be mistaken for the dark-green-brownish
jelly colonies
of Nostoc pruniforme
('Seepflaume')
found in the littoral of other lakes.
The ecosystem nutrient status can be
assessed in more detail by the allocation pattern and stoichiometric
proportion among fractions of a
source, as opposed to
the total phosphorus vs chlorophyll-a relationship. Such assessment by
triple ratios is described for the case study Old Danube. Here the
effects
of
reduced total size of phosphorus concentrations from 2.4
to 0.6 mmol L-1 on the plankton community in Old
Danube were evaluated from two triple
ratios. The relative proportions of dissolved reactive
phosphorus (DRP), dissolved non-reactive
phosphorus (DOP: mainly dissolved organic P, and some inorganic
polyphosphates) and particulate organic phosphorus (POP; mainly the
phosphorus incorporated by micro-biota drifting in the water) refers to
the three fractions of total
phosphorus. The partitioning
among the fractions of total
phosphorus is displayed by the DRP:DOP:POP
triple ratio (Fig.4a-d
in Teubner et al.
2003 R).
Secondly, the stoichiometry
among three main nutrient
elements incorporated by biota, refers to the particulate
organic
carbon (POC), nitrogen (PON), and POP, and is displayed by the POC:PON:POP triple ratio (Fig.5e
in Teubner et al.
2003 R)
.
In both triple ratios, POP was used as a key component to indicate
shifts between phosphorus fractions and between nutrient elements of
plankton biota.
At reduced phosphorus availability due to RIPLOX-treatment,
the planktonic assemblage accumulated 14% more phosphorus in their
cells than under phosphor-rich water conditions before treatment. This enhanced phosphorus accumulation by
biota was achieved in two stages. In the first year, it
was
mainly at the expense of DOP and in the second year by DRP (Fig.4b-c
in Teubner et al.
2003 R).
The
evidence indicates that this increase of %POP of TP reflects the
adaptive
phosphorus utilization of the organisms under low phosphorus supply,
substantiated by significant relationships between phosphorus fractions
and changes within the community. A higher %POP of TP was associated
with higher specific enzyme
activity (alkaline phosphatase) and with tighter
coupling between producers and
consumers respectively — that is, driven from both the
nutrient-producer and the
producer-consumer interface (Fig.4e-f
and Fig.5a-d
in Teubner et al.
2003 R).
A lowered %DOP of total
phosphorus was significantly related to smaller overall loss rates for
the carbon pool of bacteria and algae, whereas a lower %DRP of total
phosphorus was correlated with the decrease of the biomass of most
organism groups.
Stoichiometric shifts
toward a phosphorus-rich assemblage under low TP have been
justified by compositional shifts of the plankton community, namely
from nitrogen-rich cyanobacteria to phosphorus-rich eukaryotic algae
and bacteria in Old Danube (Fig.5e
in Teubner et al.
2003 R).
The stoichiometric shift toward the limiting element seems to be a common phenomenon of
individual
adaptation of producer organisms. Studies from other shallow lakes show
that
nitrogen-rich
plankton, i.e. mainly composed by cyanobacteria, dominate usually under
nitrogen-limitation (Figs.1&2
in Teubner et al.
1999 R,
Fig.4a
in Teubner & Dokulil
2000 R).
In turn phosphorus-rich
organisms, i.e. mainly
diatoms and other eukaryotic algae, dominate under phosphorus limitation
(Fig.5 f
in Teubner et al.
2003 R).
The elemental composition of individual cells of bacteria,
cyanobacteria and algae can vary strongly in contrast to consumers
keeping their elemental composition within a limited range, even if the
elemental composition of ingested food is very different from their
body tissue.
In conclusion, the shifts toward POP in both triple ratios
strongly indicated that the plankton
community acted as a sink for phosphorus under reduced total pool size
in Old Danube (abstract
& page 1148
in Teubner et al.
2003 R).
 >
>