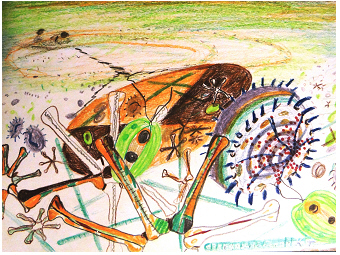
 Bildtitel: „Ein
See aus der Perspektive der
Algen“, gemalt von Katrin
Teubner, 2005:
Bildtitel: „Ein
See aus der Perspektive der
Algen“, gemalt von Katrin
Teubner, 2005:
Die mikroskopische Ansicht der Algen steht im
Vordergrund des Bildes. Hinter den Algen ist eine Secchi-Scheibe zur
Messung der Sichttiefe (siehe auch Foto unten) während einer Probenahme
zu sehen. Linksseitig oben lassen sich zwei Menschen auf dem
Probenahmeboot an der Oberfläche des Sees erkennen, die gerade das Seil
von der Secchi-Scheibe in das Wasser hinab lassen.
Wie auf dem Bild
liegt der Schwerpunkt dieser Webseite auf den photosynthetischen
Mikroorganismen, wie Algen und Cyanobak- terien. Diese mikrobiellen
Primärproduzenten spielen eine Schlüsselrolle in Seen und Flüssen.
Folgenden Fragen wird hier u.a. nachgegangen:
Wie können photosynthetische
Mikroorganismen kleine kurzfristige Nährstoffgaben nutzen?
Warum kann ein großer
Nährstoffeintrag (Eutrophierung) die Farbe des Gewässers schnell
ändern?
Warum kann eine Seenrestaurierung
durchaus scheitern bzw. warum ist die Sanierung meist langwierig und
dauert Jahre?
Woran können wir bei einem
Spaziergang am Ufer eines Sees leicht erkennen, dass an der
Wasseroberfläche Cyanobakterien zu sehen sind?
Wie lassen sich die Auswirkungen
der Klimaänderung auf Seen und speziell die im Wasser lebenden
photosynthetischen Mikroorganismen abschätzen?
Wie kann der Zustand natürlicher
und künstlicher Binnengewässer anhand der mikrobiellen Gemeinschaft von
Algen und Cyanobakterien beurteilt werden?
Wie können Mikroben und Algen
genutzt werden, um auf natürliche Weise Schwimmteiche „rein“ zu
halten
und was muss im Wesentlichen beachtet werden, um den guten Zustand
eines Schwimmteiches auf lange Sicht zu erhalten?Seen
und Flüsse sind nicht nur
Becken, in denen sich das Wasser ansammelt oder Korridore in denen das
Wasser durch die Landschaft hindurchfließt - sie sind ganz sicher viel
mehr. Wasser ist Leben! Man kann sogar sagen, dass das Wasser in
den
Seen und Flüssen selbst lebt! Natürliche Wasserbecken
können wertvolle
Lebensräume für viele Organismen sein, von einer überwältigend großen
Anzahl von mikroskopisch kleinen Lebewesen bis hin zu einer Vielfalt
von Wasserpflanzen und Wassertieren. Wir erfreuen uns an seltenen
Pflanzen und Tieren bei einer Bootsfahrt auf dem Wasser, sei es die
tropisch anmutende Wasseraloe (Stratiotes
aloides), der spektakulär
anzuschauende Geweih- Süsswasserschwamm (Spongilla
lacustris) oder der exotisch aussehende Eisvogel (Alcedo
atthis) der kleinen schilfreichen Seen und Flüsse im
Norden der
gemäßigten Klimazone. Die Begegnung mit springenden Schweinswalen (Neophocaena
phocaenoides)
in ihrem natürlichen Lebensraum der großen subtropischen Seen ist
wiederum ein besonderes Naturerlebnis. Manch ein Naturfreund ist
begeistert, solche
Habitate behutsam zu erkunden. Wir erinnern uns an Teiche, Seen, Bäche
oder Flüsse, die uns bereits in der Kindheit angezogen haben oder an
denen wir heute gern bei einem Spaziergang verweilen. Das Anlegen eines
kleinen Teiches im nahe gelegenen Wald oder in unserem Garten hat
wiederum seine eigene Faszination. All diese natürlichen und
künstlichen
Gewässer sind Teil des Lebens der Menschen. Sie sind
weltweit eng mit deren Tradition und Kultur verbunden.
Viele natürlich belassene Binnengewässer sind wertvolle Quellen für
Nahrung und Trinkwasser. Andere Binnengewässer sind stark von
Konstruktionen modifiziert und dienen der Schifffahrt oder als
Talsperre dem Hochwasserschutz, der Trinkwasserentnahme und der
Energiegewinnung. Umweltverschmutzung von Binnengewässern ist ein
weiteres Zeichen des menschlichen Einflusses auf diese Ökosysteme und
hat auch viele Aspekte. Ein Aspekt ist die Eutrophierung, wo ein
künstlich hoher Eintrag der
wichtigsten Nährelemente wie Phosphor und
Stickstoff das aquatische Ökosystem beeinflusst. Die
Konzentrationen von Nährstoffen in einem eutrophierten See oder Fluss
sind daher viel höher als jene, die dem natürlichen Hintergrund des
jeweiligen Gewässers entsprechen. Hohe Nährstoffbelastungen z.B. durch
unbehandelte
Abwassereinleitung erzeugen somit „extreme“ Umweltbedingungen. Solch
eine Nährstoffanreicherung ist häufig mit einer Verringerung der
Transparenz des Wassers verbunden. Dadurch erscheint das
getrübte Wasser intensiv
gefärbt in den Tönen
gelb-grün, dunkel-grün oder auch Bordeaux-rot, je nachdem,
welche Algen
oder Cyanobakterien sich am meisten im Zuge des erhöhten
Nährstoffeintrages entwickeln. Seen und Flüsse sind dann weit von einem
„gesunden Ökosystem“, d.h. von einem guten oder sehr guten ökologischen
Zustand (siehe alpine Seen wie
Attersee S und
Traunsee S),
entfernt. In
den meisten Fällen kann der Grund der
Nährstoffzunahme eines Sees oder Flusses nicht in dem See oder dem
Fluss selbst gefunden werden, sondern liegt in seinem oft großflächigen
Einzugsgebiet. Eine Restaurierung im Gewässer allein ist daher nicht
ausreichend , sondern muss
von einem erfolgreichen
Sanierungsmanagement
im Einzugsgebiet begleitet werden (siehe hier see-interne Sanierung der
urbanen Alten
Donau S).
Die Auswirkung von großen Nährstoffmengen wurde über eine Vielzahl von
sogenannten Dünge-Experimenten, die auch als Auswuchs-Experimente
oder Nährstoff-Zugabe-Experimente bezeichnet werden, gut abgeschätzt.
Mit solchen Bioassay-Experimenten konnten wesentliche Zusammenhänge
über
die OPTIMIERUNG der Wachstumsbedingungen von Algen im Gewässer erlangt
werden. Dieses Wissen half die Problematik der massiven Eutrophierung
grundlegend zu verstehen. Die Eutrophierung
von Binnengewässern ist ein
weltweit verbreitetes Phänomen in urbanen Regionen. Die
gesundheitlichen Risiken durch z.T. aufrahmende, Toxine bildende
Cyanobakterien wie Microcystis
spp.,
Planktothrix rubescens,
Cylindospermopsis raciborskii,
Aphanizomenon spp.
and Anabaena spp.
sind aus einer Vielzahl von nährstoffreichen Gewässern der
Länder
rund um den Globus bekannt (zu den Massenentwicklungen dieser
Cyanobakterien siehe hier Bergknappweiher
S,
Dianchi S,
Grosser Mueggelsee S,
Taihu S,
Alte Donau S,
Ammersee S und
Mondsee S). Begleitende
Untersuchungen von
Seen und Flüssen
sowie Restaurationsprogramme sind daher in vielen Ländern initiiert
worden, um die Wasserqualität zu kontrollieren bzw. zu verbessern.
Einige der auf
dieser Webseite gezeigten eutrophierten Gewässer sind vor Jahren
aufgenommen, spiegeln aber selbst heute noch diesen nicht
zufriedenstellenden Gewässerzustand dar. Andere Fotografien
mit Aufrahmungen an der Wasseroberfläche
können hier jedoch bereits eine historische Ansicht sein, da eine
Gewässersanierung
inzwischen erfolgreich war. Viel wurde bereits getan,
um Meilensteine einer grundlegenden Restaurierung vieler Seen und
Flüsse weltweit zu erreichen. Eine durchgreifende Restaurierung von
stark eutrophierten Binnengewässern konnte dabei häufig durch die
drastische Reduktion des Nährstoffeintrages aus gewässer-externen und
auch gewässer-internen Quellen erreicht werden. Der Erfolg dieser
Maßnahmen lässt sich gewässerkundlich heute einfach und plausibel
nachvollziehen, auch wenn die Kosten solcher Maßnahmen im Gewässer und
in dessen Einzugsgebiet in Summe außerordentlich hoch sind.
 „Secchi-Scheibe
(Secchi-Disk)“ am Seeufer;
eingefügtes Bild:
Secchi-Scheibe und Lichtmessgerät im Labor:
„Secchi-Scheibe
(Secchi-Disk)“ am Seeufer;
eingefügtes Bild:
Secchi-Scheibe und Lichtmessgerät im Labor:
Das Wachstum der photosynthetischen Mikroorganismen in der Wassersäule
hängt von den
Lichtverhältnissen unter Wasser ab. Die Abschätzung des
Unterwasser-Lichtklimas ist daher für die Beurteilung der Produktivität
eines Binnengewässers wichtig. Mit einer Secchi-Scheibe können
die Lichtverhältnisse unter Wasser oder die Transparenz des
Wassers am
einfachsten abgeschätzt werden. Diese Scheibe wird an einer Leine in
das tiefe Wasser herab gelassen, bis sie eine kurze Weile nicht mehr zu
sehen ist. Beim Hochziehen der Secchi-Scheibe zur Wasseroberfläche wird
dann genau die Wassertiefe gemessen, in der die Scheibe zum ersten Mal
wieder zu sehen ist. Diese Tiefe wird als Sichttiefe bezeichnet. Für
den alpinen Mondsee beträgt das Jahresmittel der Sichttiefe
3,3 m. Die
mit der Gewässertiefe abnehmende Lichtintensität kann weiters mit einem
Lichtmessgerät gemessen werden. Im Jahresmittel ist im Mondsee in einer
Tiefe von 11 m noch 1% der an der Wasseroberfläche
eintreffenden
Lichtintensität für die Photosynthese verfügbar. Diese durchleuchtete
Schicht nennt man „euphotische Zone“. In dieser Schicht überwiegt die
mikrobielle Photosynthese gegenüber der mikrobiellen Respiration (siehe
weitere Tiefen mit einem ausgeprägten Wachstum der photosynthetischen
Mikroorganismen bei 10-12% bzw. 0,1% Lichtintensität im Mondsee S
und
Ammersee S)
auf dieser Webseite). In einem See korrespondieren die Mächtigkeit der
euphotischen Zone
und die Sichttiefe miteinander. Die euphotische Zone
kann daher
gut über die Sichttiefe, d.h. einfach über eine Messung mit einer
Secchi-Scheibe,
abgeschätzt werden. Während der Vegetationsperiode vom späten Frühjahr
bis frühen Herbst reicht im Mondsee die euphotische Zone im
Durchschnitt um das 3,42-fache tiefer als die Sichttiefe. Dieser Faktor
variiert nur leicht zwischen den Jahreszeiten, von Jahr zu Jahr in
einem
See bzw. zwischen verschiedenen Seen desselben Seentypes.
In anderen Binnengewässern gestaltet sich die
Kontrolle des Wachstums der Cyanobakterien und anderer Algenblüten
jedoch komplexer. Das erneute „unerwartete“ Aufkommen solcher
Massenentwicklungen setzt häufig nach einer vor Jahren durchaus
erfolgreich durchgeführten Gewässersanierung ein. Daher sollte die
aktuelle Gewässersituation neu hinterfragt werden. Im Gewässer liegt
ein prinzipiell anderes Bedingungsgefüge als zu jener Zeit eines
übermäßig (!) hohen Nährstoffniveaus (Nährstoff-Überfluss) vor, wie es
für die Periode der massiven Eutrophierung im Abschnitt zuvor
beschrieben wurde. Das massenhafte
Neu-Auftreten von photosynthetischen
Mikroorganismen tritt nämlich zu einem Zeitpunkt auf, zu
dem
„eigentlich“ bereits ein Nährstoff-Mangel (!) im Gewässer herrscht.
Es konnte durch vielfältige Gewässeruntersuchungen gut belegt werden,
dass in Gewässern mit Nährstoffmangel die Umbauzeiten von Nährstoffen
schneller geschehen und auch vielfältigere Wege gehen als in Gewässern
mit Nährstoff-Überfluss. Diese Mechanismen können über die Abschätzung
von Stoffkreisläufen aber auch den Grad
der Kopplungen zwischen den
Produzenten, d.h. den im Wasser schwebenden Algen und
Cyanobakterien
(Lebensgemeinschaft Phytoplankton), und den Konsumenten, d.h.
beispielsweise den kleinen Tieren der Gemeinschaft des Zooplanktons,
aufgezeigt werden (siehe z.B. Abb.
5
A und B
in Teubner et al.
2003 R).
Die
Nährstoffarmut im Gewässer
wird durch eine erhöhte AKTIVITÄT der
ORGANISMEN der mikrobiellen Gemeinschaften beantwortet,
wodurch die
wachstumsbegrenzenden Nährstoffe rasch wieder freigesetzt und somit
innerhalb kurzer Zeitspannen etwa dem Wachstum der Algen erneut zur
Verfügung stehen (u.a. erhöhte
mikrobielle Aktivität durch die Bildung extrazellulärerer Enzyme wie
Phosphatasen bei Phosphor-Armut im Gewässer, siehe Abb. 4
F
in Teubner et al.
2003 R).
Diese Regulationsmechanismen, die es erlauben mit
einem
Nährstoffmangel gut umgehen zu können, spiegeln sich schlussendlich
beispielsweise in dem Neuaufkommen der Cyanobakterien wieder. Mit einem
tieferen Verständnis kann man sogar sagen, dass diese
Regulationsmechanismen auf der
Fähigkeit der Organismen, unmittelbar
auf ihre momentane Umweltsituation zu reagieren, begründet
liegen. Die
heutige Sichtweise in der Gewässerökologie konzentriert sich zu sehr
auf langfristige Veränderungen der mikrobiellen Gemeinschaft über
mehrere Generationen (z.B. Ermittlung der Veränderung der
Artenzusammensetzung der mikrobiellen Gemeinschaften in
Bioassay-Experimenten), wodurch ein wesentlicher Blick eines
Gewässerbiologen, nämlich auf die ORGANISMEN im Ökosystem, verstellt
wird. Es kann davon ausgegangen werden, dass mikrobielle
Primärproduzenten,
wie Algen und Cyanobakterien, nur einen Tag oder einige wenige Tage
unter günstigen Wachstumsbedingungen in einem Gewässer leben, bevor sie
eine Zellteilung durchlaufen. Wie können sich diese Mikroorganismen auf
unvorhersehbare oder vorhersehbare Änderungen ihrer Umwelt innerhalb
ihrer kurzen Lebensdauer von nur einem Tag-Nacht-Zyklus einstellen?
Diese andere Perspektive der Interaktion von Mikroorganismen mit ihrer
Umwelt in biologisch
relevanten, räumlich und zeitlich kleinen Skalen,
hilft verstehen, wie die Ausnützung
von kurzlebigen Nährstoffquellen
durch Algen in nährstoffarmen Gewässern geschieht. Dies
soll nun im folgenden etwas näher für die Aufnahme des Nährstoffes
Phosphat in den nachfolgenden zwei Abschnitten beschrieben werden.
Die Phosphataufnahme erfolgt nach einem völlig anderen Prinzip als die
gut erforschte, sättigende Substrataufnahme der Enzymkinetik. Dem
häufigen Szenario des
Phosphatmangels
in natürlichen Binnengewässern
Rechnung tragend, ist bei Algen kein sättigendes
Phosphat-Aufnahmesystem ausgebildet, das eine Inkorporation dieses
Nährstoffes etwa abbricht. Es wird quasi so viel wie möglich an
Phosphat
aufgenommen. Bei hohen Phosphatgaben können die Mikroben über den
sogenannten „Luxuskonsum“ große Mengen an Phosphat aufnehmen. Die in
den Zellen gebildeten Phosphat-Granula sind dann mikroskopisch leicht
zu erkennen. Im Extremfall der unbegrenzt hohen Phosphatzugabe, wie
z.B. bei Experimenten, nehmen die Zellen eine sehr große Menge, nämlich
das Vielfache ihres üblichen zellulären Phosphatgehaltes auf. Nach
einer solch übermäßigen Phosphataufnahme sind diese Mikroorganismen
jedoch nicht weiter lebensfähig. Wie zuvor aber bereits erwähnt, sind
natürlicher Weise extrem hohe Phosphatkonzentrationsgaben in
Binnengewässern nicht wirklich zu erwarten. Der starke Mangel an
Phosphat ist die häufigere Situation, an die sich die
photosynthetischen Mikroorganismen mit ihrem
adaptiven Aufnahmesystem
angepasst haben. Neuere Studien zeigen, dass Algen sehr wohl in der
Lage sind, durch rasche chemotaktische und kinetische Reaktionen kleine
Punktquellen von Nährstoffkonzentrationen ausnützen zu können, bevor
diese durch physikalische Prozesse wie Diffusion im Wasserkörper
verteilt werden. Solche kurzlebigen, punktförmigen Nährstoffquellen
entstehen z.B. durch individuelle Exkretionen einer Vielzahl von Tieren
im Wasser, von den Einzellern (Protozoa) bis hin zu den Fischen. Das
Konzentrationsniveau, bei dem die Algen und Cyanobakterien durchaus
noch effizient Phosphat aufnehmen können, liegt unterhalb der
chemischen Nachweisgrenze (Nachweis bis zu 140 nanomol
pro Liter im
Süßwasser nach Standardmethoden der Gewässerchemie für Süßwasser;
Teubner et al., Publikation in Vorbereitung).
Daher können die
Primärproduzenten selbst in solchen Gewässern noch erfolgreich wachsen,
in denen nach Standardmethoden kein freies Phosphat mehr nachgewiesen
werden kann.
Das Phosphat kann nur über einen
aktiven Transport, angetrieben durch
metabolische Energie, von den photosynthetischen Mikroorganismen
aufgenommen werden. Die P-Aufnahmesysteme folgen dabei einem
„steady-state“, besser gesagt einem stationären Zustand der
Nichtgleichgewichtsthermodynamik. Bei der Aufnahme von portionsweise
kleinen, d.h. nanomolaren, Phosphatmengen, wie sie z.B. durch
Ausscheidungen von Zooplankton den Algen verfügbar sind und nachfolgend
diskutiert werden, ergeben sich nur geringe Abweichungen von diesem
thermodynamischen Gleichgewichtszustand. Unter diesen Bedingungen
erfolgt die Phosphataufnahme über eine lineare Abhängigkeit einer
thermodynamischen Fluß-Kraft Beziehung, die das Aufnahmesystem immer
wieder in einen stabilen steady-state Zustand zurückführt. Solch ein
aktives Phosphat-Aufnahmesystem ist an eine Produktion von Entropie
gekoppelt
(Entropieproduktion > 0, siehe z.B. Glaser 2005). Nach
Prigogine streben solche Systeme eine minimale Entropieproduktion an
(Prigogine’s Minimum der
Entropieproduktion, Glansdorff & Prigogine 1971).
Es mag hier
ergänzt werden, dass damit die Phosphataufnahme nach einem anderen
thermodynamischen Prinzip verläuft als der Aufbau dissipativer
Strukturen wie Wachstum oder Fitness, bei denen die Systeme aufgrund
der Bildung neuer Strukturen auf eine maximale Entropieproduktion
abzielen. Die Aufnahme von Phosphat ist nicht zwangsläufig an eine
Zellteilung, und damit auch nicht an Wachstum gekoppelt
(Zweikompartment-Modell, siehe Droop 1973). Die
Entropieproduktion während der Aufnahme mehrerer kleiner
Phosphatgaben lässt sich empirisch gut über die Kenngrößen einer
Fluß-Kraft Beziehung bestimmen. Die Optimierung
der Phosphataufnahme bei Algen
kann damit phänomenologisch nach dem Ausmaß der daran gekoppelten
Entropieproduktion betrachtet werden. Die Entropieproduktion wird dabei
als Aufwand der Phosphataufnahme durch die Algen interpretiert. Es
zeigt sich, dass es für die Algen von Vorteil ist, eine bestimmte Menge
Phosphat in kleinen nanomolaren Portionen aufzunehmen, da hier die
Entropieproduktion relativ niedrig ist, d.h. das Phosphat mit relativ
wenig Aufwand aufgenommen werden kann (Teubner et al., Publikation in
Vorbereitung). Bei einer Gabe der gleichen Menge an Phosphat aber in
Form von wenigen größeren nanomolaren Portionen ist die
Entropieproduktion vergleichsweise höher. Man sollte bedenken, dass in
natürlichen Binnengewässern, die wenig durch den Menschen beeinflusst
sind, kleine Nährstoffgaben ohne Zweifel viel häufiger vorkommen als
große. Die Individuenzahlen für einzelne Gruppen von kleinen Tieren im
Wasser schwanken in etwa zwischen 1000 und 1000 000 Individuen pro
Liter (Teubner et al., Publikation in Vorbereitung). Vier Gruppen, die
alle
der Lebensgemeinschaft Zooplankton angehören, sollen hier beispielhaft
genannt werden: die sich tierisch ernährenden einzelligen Geißeltiere,
die Wimperntierchen, die Rädertierchen und die Wasserflöhe. Durch die
Ausscheidung dieser kleinen Tiere werden punktuell jeweils nur geringe
Phosphatmengen freigesetzt. Weit seltener kommen größere Tiere, wie z.
B. Jungfische vor, wo die Ausscheidung eines einzelnen Tieres einen
wesentlich höheren Nährstoffpuls darstellt als bei den zuvor genannten
kleinen Tieren. Man kann in etwa abschätzen, dass in 1000 Liter nur ein
Jungfisch vorkommt und die Zahl der ausgewachsenen Fische nochmals weit
niedriger ist. Die Algen können sich mit ihrem aktiven
Phosphataufnahmesystem leicht darauf einstellen, ob sie sich gerade in
der häufigen Umgebung von vielen kleinen Phosphatgaben inmitten einer
„Wolke“
eines Zooplanktonschwarmes oder in der weit seltener vorkommenden
Umweltsituation mit großen Phosphatgaben inmitten eines Fischschwarmes
befinden. Im Verlauf der Aufnahme vieler kleiner Phosphatgaben können
die Algen auch von einem rasch aufnehmenden Modus in einen
zweiten, weniger aktiven Aufnahmemodus abrupt wechseln. Die
Anpassungsstrategien der Algen
bei der Phosphataufnahme sind weit
vielfältiger als es hier nur kurz angeführt wurde (Teubner et al.,
Publikation
in Vorbereitung). Sie basieren auf den interaktiven Wechselwirkungen
zwischen den Mikroorganismen und ihrer Umwelt.
Über die Dynamik in zeitlich
kurzen und räumlich kleinen Skalen, wie
sie zuvor kurz beispielhaft erörtert wurden, ist derzeit nur wenig
bekannt, obwohl diese Ökosystem-Kenntnis wichtig wäre, um die
Entwicklungsbedingungen der kurzlebigen Mikroorganismen in ihren wenig
weit ausschweifenden Lebensräumen besser verstehen zu können. Somit
würde sich auch das „unerwartete“ Neuaufkommen von problematischen
Massenentwicklungen besser erklären lassen. Neu auftretende
Massenentwicklungen von aquatischen Mikroorganismen in Gewässern mit
Nährstoffmangel sind wiederum nicht zu unterschätzen, da sie meist mit
einer erneuten Beeinträchtigung der Gewässernutzung einhergehen. Ein
gefärbtes und unangenehm riechendes Oberflächenwasser kann weder als
Trinkwasser genutzt werden, noch genügt es den hohen Ansprüchen eines
attraktiven Erholungsortes in touristischen Gebieten. Selbst in
künstlichen Gewässern, wie den natürlich gestalteten Schwimmteichen,
können unerwünschte Entwicklungen von Algen und Cyanobakterien auf
einem relativ geringen Nährstoffniveau auftreten, sofern der Teich
nicht nachhaltig angelegt wurde. In kleinen Schwimmteichen werden
„unerwartete“ Algenentwicklungen häufig viel schneller als in
natürlichen Seen festgestellt, nämlich bereits in der ersten Saison in
kleinen Becken bzw. nach einer Laufzeit von nur 3-5 Jahren in größeren
Schwimmteichanlagen (siehe naturnah
gestaltete Teiche S).
Mit anderen
Worten: Künstliche Wasserbecken
reagieren meist viel sensitiver auf geringfügige Nährstoffeinträge als
natürliche Ökosysteme. Solche Themen der Nährstoffproblematik sowie
weitere Aspekte zur Entwicklung von Primärproduzenten in
Binnengewässern werden auf dieser Webseite für verschiedene
Gewässertypen aufgezeigt. Die Schlüsselworte, die auf der Seite zu den
Publikationen in der rechten Spalte S
aufgelistet sind, geben dabei einen
Überblick, welche Themenbereiche insbesondere auf dieser
Seen-Fluß-Webseite erörtert werden.





 >
>